E v o l o c u s
The Journal of Experimental Neuroevolution, published by Evolocus LLC, ISSN 2164-6481
Vyssotski M. Action acceptor in evolution. Evolocus 1, 37-53 (2024).
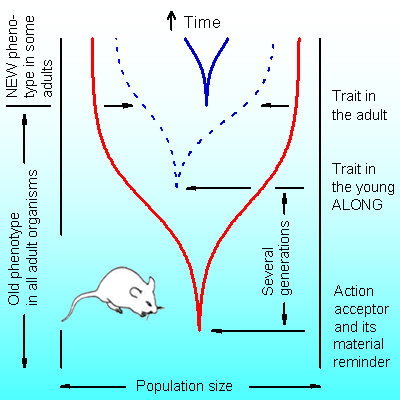
Abstract. The desired result of any process is represented in an organism by an action acceptor, consisting of two parts: the part which is absent yet and the part that is represented by material reminders of the said action acceptor. The absent part belongs to ideation space. There is no physical time in the ideation space: all entities are given simultaneously. The entities comprise, in general, a computably non-enumerable set – one entity can contain in itself one or more the same or different entities (no conservation laws). This feature provides the possibility to resolve contradictions that are unsolvable in 3D physical space. Evolution proceeds through the interaction of ideation space with 3D physical space by means of action acceptors. The development of each action acceptor precedes the appearance of its positive result by several generations and consists of selection of its material reminders through the self-election of organisms for differential reproduction.
Supplementary Audio MP3 for Vyssotski, 2024.
Supplementary Audio WAV for Vyssotski, 2024.
_________________________
Vyssotski D.L. Hybrid vigour and hybrid dysgenesis. Evolocus 1, 31-36 (2019).
Abstract. The formation of a hybrid phenotype in mammals remains a mystery. In order to resolve this mystery we have chosen inbred mouse strains C57BL/6J and DBA/2J, and their F1 hybrid B6D2F1, took only females for experiment and housed them during postnatal days P22-P60 either in standard or enriched living conditions, always 4 mice per cage. This adolescent cage enrichment has evoked either induction of hybrid vigour or its significant enhancement, observed in different operant behavioural tasks during the rest of their life (Morris water maze, Go/NoGo sound frequency and sound duration discrimination tasks). This induction or enhancement can be understood only if ontogenesis is an active process, driven by action acceptors – by entities that percept the achievement of positive developmental results, even if the last ones have appeared with a help of random or unexpected factors of stochastic or genetic nature. The same way action acceptors direct evolution on Earth.
Supplementary Information for Vyssotski, 2019.
Full Text + SI (Combined PDF)
_________________________
Neurologger 3 and its history. Evolocus 1, I-VI (2016).

Abstract. Neurologger has been originally designed to record EEG, local field potentials and neuronal activity in freely moving animals in their natural environments. Later its spectrum of applicability was extended to EMG and ECG recording. Recently Neurologger has been applied to study of auditory communication in animals and, thanks to increased up to 200 kHz sampling rate in single-channel mode, to study of ultrasonic echolocation in bats. Today Neurologger is represented by two versions: Neurologger 2A/2B and Neurologger 3. Neurologger 2A/2B remains our lightest version (1.3 g without battery) and it is capable to record up to 4 channels at 33.3 kHz, 10-bit into soldered on board memory (1 or 2 GB). Neurologger 3 is slightly heavier (1.7 g without battery), it records up to 32 channels at 20.8 kHz, 16-bit into microSDXC card (4-256 GB) and it has remote control and data access through BlueTooth communication with Windows 10 machines.
Supplementary Information for "Neurologger 3 and its history", 2016.
Full Text + SI (Combined PDF)
_________________________
Vyssotski D.L. Nomogenesis and the logic of chance. Evolocus 1, 25-30 (2016).
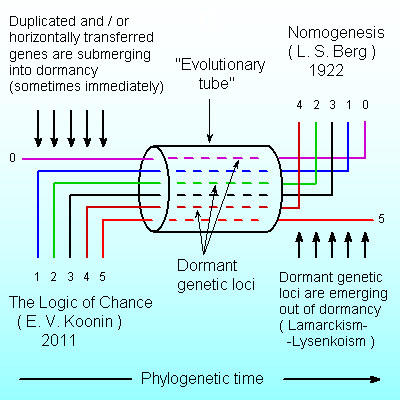
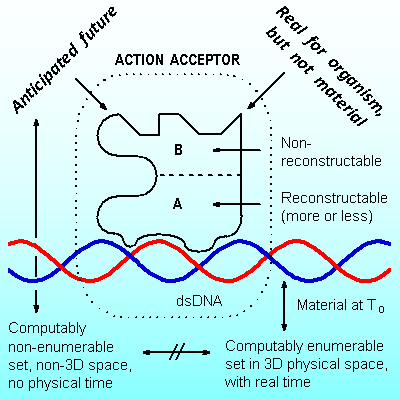
Abstract. Evolution of available genomes was shown to be proceeding through random changes, the changes that comprise the main modus of evolution (Koonin, 2011). Morphological evolution of available and extinct Metazoa was shown to be going on the basis of law, by means of precession of characters, where characters originally manifested in the young along in the course of time and evolution were displayed also in adult descendants (or supposed descendants) of that organism (Berg, 1922). This contradiction is obviously solved in nature, where the appearance of any new genetic locus in the genome and its further expression in the phenotype can be separated by unlimited period of time and by unlimited number of generations. The management of dormant genetic loci has come from the previous evolutionary stage, unimaginable today, where organisms were open systems with respect to the flow of genetic elements and were collecting, discriminating and storing genetic elements from the external environment. This was an important period when multiple systems for blocking and unblocking of genetic loci came into being. However even before this stage, it was even more fantastic evolutionary period where replication, transcription and translation were absent and Eigen cycle was not possible, but organisms were collecting randomly available components (proteins, RNA and DNA) by means of action acceptors (Anokhin, 1955) – sites of double-stranded DNA mechanically compatible with useful components. Action acceptors themselves were unable to be replicated by modern way (no DNA polymerase!), but they were collecting their pseudo-copies from the environment – the pieces of DNA that were born in the environment and occurred to be compatible by chance with current action acceptors. Action acceptors, – the structures that sense presumably useful results or substances, were directing evolution from the early beginning and they are directing it today through activation and deactivation of dormant genetic loci.
Supplementary Information for Vyssotski, 2016.
Full Text + SI (Combined PDF)
_________________________
Vyssotski D.L. Transgenerational epigenetic compensation and natural selection. Evolocus 1, 19-24 (2014).
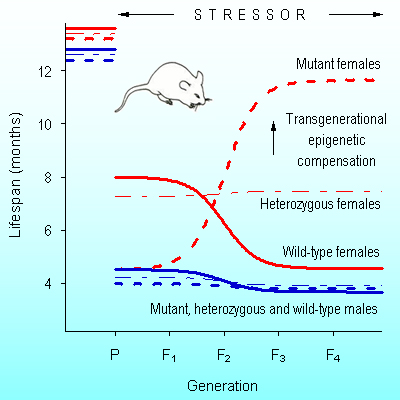
Abstract. The term “epigenetics” defines all meiotically and mitotically heritable changes in gene expression that are not coded in the DNA sequence itself. Transgenerational epigenetic compensation was discovered in the untreated progeny of drug-treated males (rats and mice) as the opposite quantitative phenotypic changes. In natural populations, the hereditary basis of transgenerational epigenetic compensation develops mainly in homozygous mutant males, but it does not affect their phenotype. In their descendants, being in an independent locus, this heritable epigenetic compensation increases fitness and lifespan of homozygous mutant females and decreases lifespan of wild-type females, starting from F2. Here we show that this transgenerational epigenetic compensation is a guiding agent of natural selection. Natural selection is not a directing or driving force of evolution anymore. Natural selection needs some guidance. Transgenerational epigenetic compensation can initiate speciation through segregation of mutants and wild-types and/or it can change the selection coefficient of a given mutation.
Supplementary Information for Vyssotski, 2014.
Full Text + SI (Combined PDF)
_________________________
Vyssotski D.L. Transgenerational epigenetic compensation and sexual dimorphism. Evolocus 1, 13-18 (2013).
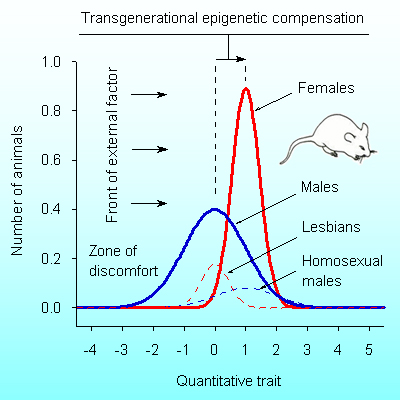
Abstract. The term “epigenetics” defines all meiotically and mitotically heritable changes in gene expression that are not coded in the DNA sequence itself. Transgenerational epigenetic compensation was discovered in the untreated progeny of drug-treated males (rats and mice) as the opposite quantitative phenotypic changes. In natural populations, heritable epigenetic compensation can convert mutants into hopeful monsters, initiates speciation, and therefore determines the route of macroevolution. Transgenerational epigenetic compensation facilitates genetic assimilation of acquired characters in microevolution. The ontogenesis of females is better canalized than that one of males, and natural selection proceeds mainly through selection of males. However the presence of sexual dimorphism in transgenerational epigenetic compensation remains unclear. Here we show that the hereditary basis of transgenerational epigenetic compensation develops mainly in males. However the phenotypic results of this development are more pronounced in their female descendants, starting from F2. This sexual dimorphism enhances the efficiency of micro- and macroevolution.
Supplementary Information for Vyssotski, 2013.
Full Text + SI (Combined PDF)
_________________________
Vyssotski D.L. Transgenerational epigenetic compensation in evolution. Evolocus 1, 7-12 (2012).
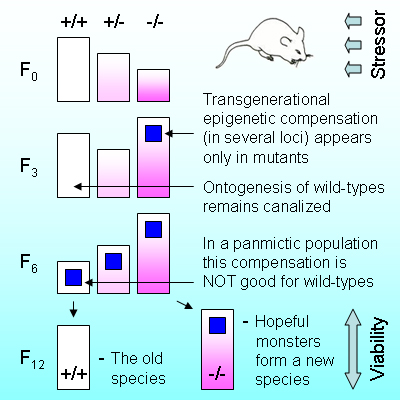
Abstract. The term “epigenetics” defines all meiotically and mitotically heritable changes in gene expression that are not coded in the DNA sequence itself. Transgenerational epigenetic compensation of disturbed functionality was discovered in the untreated progeny of drug-treated fathers as the opposite quantitative phenotypic changes (phenotypic inversion). Epigenetic changes, responsible for heritable compensation, are distributed between several independent loci and these changes disappear gradually and asynchronously during a few untreated generations. The role of hereditary epigenetic compensation in evolution remains unclear. Here we show that transgenerational epigenetic compensation of disturbed functionality converts mutants into hopeful monsters, initiates speciation and facilitates genetic assimilation of acquired characters. The increase of environmental pressure, applied to mutant and wild-type animals, induces heritable epigenetic compensation in mutants (initially less fit), whereas the development of wild-types remains canalized. In a random breeding population this heritable epigenetic compensation increases fitness and lifespan of mutants and decreases lifespan of wild-types.
Supplementary Information for Vyssotski, 2012.
Full Text + SI (Combined PDF)
_________________________
Vyssotski D.L. Transgenerational epigenetic compensation. Evolocus 1, 1-6 (2011).
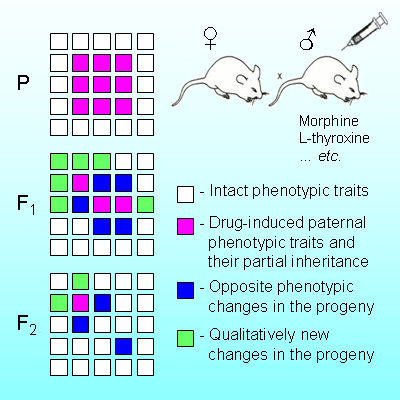
Abstract. The term “epigenetics” defines all meiotically and mitotically heritable changes in gene expression that are not coded in the DNA sequence itself. The ability of environmental factors to reprogram the germ line and to promote transgenerational disease states has significant implications for evolutionary biology. However, the biological function of transgenerational epigenetic inheritance remains unclear. Here we show that epigenetic inheritance promotes transgenerational compensation of disturbed functionality. After chronic morphine treatment of male Wistar rats or neonatal thyroxine treatment of male DBA/2J mice many of the changes discovered in the untreated progeny occurred to be the opposite of those observed in the treated fathers themselves. Phenotypic analysis of the untreated F1-F3 generations has revealed several independent epigenetically modified loci. Transgenerational epigenetic compensation was observed in the F2-F3 and further generations of transgenic Per2(Brdm1) mice raised under semi-natural outdoor conditions and it was localized not in the same locus as original mutation.
Supplementary Information for Vyssotski, 2011.
Full Text + SI (Combined PDF)
_________________________
Neuroscience 2024
Booth #1683, October 5-9, Chicago, IL

_________________________
Neuroscience 2023
Booth #2938, November 11-15, Washington, DC


_________________________
Neuroscience 2019
Booth #1764, October 20-23, Chicago, IL

_________________________
Products of Evolocus LLC

Holeboard-based novel object recognition task

This system can be used as a classic 16-hole hole-board or it can be used with any known protocol of novel object recognition task. In our task a mouse can discriminate between holes in the floor with different shapes (round, square, 6-point star, etc.) and/or between different floor materials. In addition, this system works well for any olfactory discrimination.

Technically, device is recording nosepoke activity with a help of infrared beams under the floor. Typical system consists of 4 independent chambers (about 16’’ x 16’’ each) and 4 mice can be tested simultaneously. There are two versions available. The first one is optimized for Noldus EthoVision (3.1 or any previous) and it is intended for use with Noldus video-tracking hardware and software for data acquisition and analysis. The second version is independent from Noldus EthoVision and it does not use video-tracking.

Our system has high discriminative power and it was verified using C57BL/6J, DBA/2J and B6D2F1 mice after standard and enriched housing conditions. Mice were taken from: Wolfer et al., Nature 432, 821-822 (2004). [Supplement 1, 2, 3]

Brief introduction into the history of Hole-board task
During the last 25 years Holeboard task has evolved from classic version with manual recording of mouse behavior towards modern computer-controlled system with infrared beams under the floor. Modern system can register duration of exploratory episodes to within 10 ms (using the same classic geometry with 16 holes (d = 25 mm) in the 40 × 40 cm arena, surrounded by vertical walls). In the classic Holeboard the single specific indicator of behavior was the number of nosepokes (for example, during 6-min session). In the modern setup it was discovered that two other indicators have better discriminative power: 1) “total exploration time” (this time is about 40-70 s for 6-min session) and, especially, 2) mean “nosepoke duration” (typical nosepoke duration is about 0.5-1.2 s). Contrary to straightforward logic, mean nosepoke duration can provide higher statistical significance than the number of nosepokes or total exploration time.
Video tracking is also useful for Holeboard task. It can not be used instead of nosepoke registration, but systems with simultaneous video tracking and infrared beams under the floor are available now. The best ones can video track 4 animals in 4 Holeboard units with concurrent registration of all nosepoke activity.
In addition to above-mentioned technical progress, there is significant progress in Holeboard protocols. Today after the application of a classic hole-board protocol during the first two days (for investigation of habituation) it is reasonable to apply al least one day of visual-tactile discrimination and at least one day of olfactory discrimination. During the visual-tactile discrimination test at least one hole of Holeboard is replaced by a hole with different shape [for example, circular hole is replaced by star-shaped hole], and/or the floor material, corresponding to at least one above-mentioned hole, is replaced by material which differs in color and/or surface texture; above-mentioned material can also have entirely different origin and/or production history [as, for example, plywood in comparison with polyvinyl chloride]. During olfactory discrimination test the unusual odor is placed under at least one hole of Holeboard [it can be, for example, Mint odor]. Usually during olfactory discrimination test the classic floor with 16 circular holes is used. There are publications with 4-hole Holeboard for olfactory discrimination, but the decrease in the number of holes leads to significant decrease in discriminative power (i.e. efficiency) of Holeboard test, at least in accordance with our observations. During both visual-tactile and olfactory discrimination we recommend to use equal numbers of new and old holes (e.g. 8 circular holes vs. 8 star-shaped holes and 8 holes without odor vs. 8 holes with Mint odor). Minimal modern protocol consists of 4 days: days 1 & 2 – classic Holeboard, day 3 – visual-tactile discrimination, day 4 – olfactory discrimination.